Review of Jonathan Wells’s New Book Zombie Science (Part 1)
Jonathan Wells, noted intelligent design advocate and author of the influential book Icons of Evolution, 1 has published a sequel entitled Zombie Science: More Icons of Evolution (Fig. 1). 2 In Icons, Wells showed how alleged evidences for macroevolution such as the Miller-Urey experiments, Darwin’s tree of life, industrial melanism of peppered moths, the horse fossil record, Haeckel’s embryos, Darwin’s finches, Archaeopteryx, and many others had been disproved or even exposed as frauds. The subject of his new book is how disproved evidences for macroevolution are nevertheless still being taught and hence have become Zombie Science. Wells revisits the icons he discussed in his previous book and then discusses new ones. This review will touch upon the more important points brought out in the book. Wells, who has his doctorate in molecular and cell biology, is a good writer with a keen command of his subject. He writes clearly and concisely with penetrating insight and a sense of humor.
Chapter 1: Who Let the Zombies Out?

Figure 1
Wells uses this chapter to give the reader background, define terms, and set the stage for the rest of the book. Science has a position of authority in our culture, but claims are often made that are proved incorrect and have to be retracted. Wells discusses four types of science. Science in an endeavor to discover truth through observation, experiments, and hypotheses. If a hypothesis is repeatedly found to be inconsistent with the evidence, it should be revised or rejected. This is how empirical science should work. Technological science applies what we have learned through empirical science to the betterment of human life via medicine and technology. Consensus science relies upon the prevailing opinion among scientists to determine truth and is often wrong. Past examples of consensus science gone wrong are geocentrism and spontaneous generation. The last type of science is called methodological naturalism. It is the belief that experiments can only be used to investigate physical causes. It does not assume that nonphysical causes such as intelligence don’t exist, just that experiments are not useful for studying nonphysical causes. Hence methodological naturalism acknowledges the limits of empirical science. This is different from the philosophy of metaphysical naturalism which holds that the natural world—with its mass/energy, space/time, and physical laws—is all there is; all phenomena must have a physical cause—immaterial causes such as intelligence (God) are forbidden from consideration because they do not ultimately exist. Intelligent design, on the other hand, assumes there are some features of the world that are best explained by intelligent causes, such as the coded information found in DNA.
Modern science is dominated by the materialistic philosophy. Material causes are sought despite the evidence for intelligent design. When materialistic science masquerades as empirical science, it creates Zombie Science. Zombie science is clearly seen in evolutionary biology.
Wells discusses micro- and macroevolution. Microevolution (variation within kinds) involves small variations within a population of organisms and has been repeatedly observed. Microevolution is part of empirical science. Macroevolution, molecules-to-man-evolution, extrapolates microevolution to very large changes without much empirical support and is more like a philosophy. Darwin said the evidence available to him in the biological world was consistent with descent with modification from a common ancestor. This is true for microevolution, but has not been demonstrated for macroevolution. Darwin made the extrapolation of microevolution to macroevolution thinking that future research would fill in the gaps. Darwin was wrong. Darwin’s mechanism for evolution was random variations acted upon by natural selection. He provided no details about the origin of organs and new body plans. He knew nothing of genetics or biochemistry. Eventually, genetics and Darwinism were combined into the neo-Darwinian synthesis: evolution allegedly works by random mutations in DNA acted upon by natural selection. Nowadays, microevolution is discussed as if it were macroevolution on a small scale, which is unfounded. Often, writers provide the ample evidence for microevolution and then assert the same “overwhelming evidence” is proof of macroevolution, a bait-and-switch ploy.
Chapter 2: The Tree of Life
Carl Linnaeus, founder of the science of taxonomy, believed the nested hierarchies seen in the biological world were part of divine creation. 3 Darwin believed that the similarities between different organisms were due to common ancestry. Features that were similar and thought to have derived from common ancestry were said to be homologous. Homologies are features similar in structure or position derived from common ancestry but may serve different functions. For example, a bat’s wing and the human hand are thought to have both originated from a common ancestor with an appendage with five digits. Analogies, in contrast, are features with similar structures that serve the same function but were the result of independent adaptation, not common ancestry. An example of analogous structures is the camera eye in the octopus and humans. Some scientists believe homologies are due to a common plan or design by an intelligence, not common ancestry.
Darwin believed there was an original life form from which all other life descended. Hence the pattern of similarities of organisms was predicted to be like a tree. Also predicted were innumerable intermediate forms. The intermediates which Darwin hoped would eventually be found in the fossil record are still missing. Imperfection of the fossil record is no longer a valid excuse for their absence. The Cambrian Explosion at the base of the fossil record reveals that many diverse creatures came on the scene in a short period of time without any apparent ancestors beneath. The situation has provoked evolutionists to propose “ghost lineages” (that must have existed if evolution is true). The Cambrian Explosion was an information explosion. There were many complex organisms present from the start. The fossil record is characterized by sudden appearance, stasis, and extinction and not by continuous series of slightly different forms as predicted by gradualism.
The theory of punctuated equilibrium was advanced to account for the lack of transitional forms. Hence macroevolution presumably occurred in spurts when extreme environmental changes required rapid adaptation. But this mechanism requires the same amount of information be generated as gradualism but in less time making it even less probable.
In 2014, British biologist Ronald Jenner wrote that our “imagination” is the only tool that can bridge the fragmentary fossil evidence into a coherent historical narrative. One can’t establish ancestor/descendant relationships based on fossils. Phylogenetic trees assume common ancestry but do not establish it. Designed objects can also be arranged in phylogenetic trees. Hence common features between organisms could be because of a common designer. Other methods used to compare organisms such as cladograms or molecules suffer from the same ambiguities. In addition, using different molecules for comparison can result in different phylogenetic trees. Attempts to use several molecules simultaneously have not solved the problem. How to align base sequences is not always clear-cut. Computer programs used for the alignments can make mistakes. Moreover, the phylogenies based on fossils and molecules often disagree, adding to the confusion. Another problem is that some organisms assumed to have diverged from a common ancestor long ago nevertheless have some molecules with similar sequences. This is explained by convergent evolution. In the end, evolution from a common ancestor is assumed and data in conflict with the expected pattern is often considered suspect and rejected.
Another problem is that of orphan genes. Orphan genes are only found in a single species and hence don’t appear to have been inherited. New genes are thought to evolve by gene duplication and mutation. Presumably, the duplicate gene eventually mutates into a new useful gene while the original continues to serve its function. Such a mechanism should leave evidence in the descendants. Orphan genes are not predicted by this mechanism. Orphan genes have been found wherever complete genomes have been sequenced: many bacteria, yeast, fruit flies, mice, humans, squids, and octopuses. Evolutionists claim that orphan genes evolved from noncoding, nonfunctional DNA. They propose “ghost lineages” much the same way as for fossils. Orphan genes are often ignored when constructing phylogenetic trees. This occasionally requires ignoring more than half of the genes!
Because of the difficulties above, some evolutionists say there was never a single common ancestor to life but several.
There is a developmental theory of homology. It was once believed that similar (homologous) structures would always be specified by similar genes, but this has not been borne out by experiment: similar genes don’t always produce similar structures and similar structures are not always derived from similar genes. Another theory held that similar cells during development would produce similar structures. But we now know that similar (homologous) structures can’t be traced back to cells in the same embryonic positions or to the same parts of the egg. Indeed, homologous structures are not even brought about by the same developmental mechanisms.
Circular reasoning is used when constructing phylogenetic trees. Similar structures are assumed to be the result of common ancestry, and the degree of similarity is then used to build a tree. The tree is then thought to affirm common ancestry, but common ancestry was assumed in order to build the tree.
Homologies can’t be explained by identity of genes that direct their development. Homologies are now attributed to developmental gene regulatory networks that underlie the evolution of developmental pathways. But that is still saying that DNA is responsible for homologies—very confusing.
Analogous structures are similar structures that can’t be the result of common ancestry. They are presumably the result of convergent evolution. 4 So similar structures are evidence for common ancestry except when they are not! Wells gives examples of analogous structures: echolocation (in bats and dolphins) and camera eye (in humans and octopuses). Convergence is ubiquitous. There are convergent animals such as wombats (marsupial) and ground hogs (placental). There are convergent plants. One explanation is that there are only a few preferred pathways for evolution to traverse. So since there is little chance that a random mutation/natural selection process could stumble upon the same solutions over and over again, there must only be a few ways evolution could work. And yet the variety of life on earth is enormous. If analogous structures are inevitable, does that mean nature has been preprogrammed? Sounds like design.
Chapter 3: Survival of the Fakest
In this chapter, Wells revisits some of the icons he wrote about in his first book. He revisits the Muller Urey Experiment. Urey and Miller wanted to investigate what the early earth’s chemistry may have been like. They assumed earth’s early atmosphere was reducing and conducted their experiments accordingly. The used methane, ammonia, hydrogen, and water with heat and electrical sparks. All oxygen was eliminated. They were able to produce amino acids and a few other building blocks, but produced mainly tar (which was toxic to bacteria). However, further investigation by geochemists revealed that the atmosphere of the early earth was never reducing but neutral, consisting of water vapor, carbon dioxide, and nitrogen. Experiments conducted using a neutral atmosphere produced only one simple amino acid (alanine). Methane was required to produce the other amino acids. Indeed, some of Miller’s reaction product vials did contain all 22 amino acids founds in living things (among other things), but since he started with the wrong atmosphere, the results are irrelevant to abiogenesis. Nevertheless, The Miller-Urey experiments are still taught as showing how the basic building blocks of life could have formed on the ancient earth. None of the alleged steps in the origin of life stories have been empirically demonstrated.
Then there were Haeckel’s embryos. Haeckel was a contemporary of Darwin. Darwin believed that the similarities of the early stages of embryos of various organisms were important evidence for his theory. Haeckel made draw- ings of the early stages of developing embryos of various organisms (Fig. 2). The drawings showed great similarity implying they all had a common ancestor and were going through similar early phases of development. The expression “ontology recapitulates phylogeny”—meaning the stages of embryonic development mirror the organism’s evolutionary history—applies to this thinking. However, it turned out that the drawings were faked! In reality, the most similar stages of the developing embryos comes much later in development; the early stages are more dissimilar. Nevertheless, Haeckel’s embryos are still taught in biology textbooks.
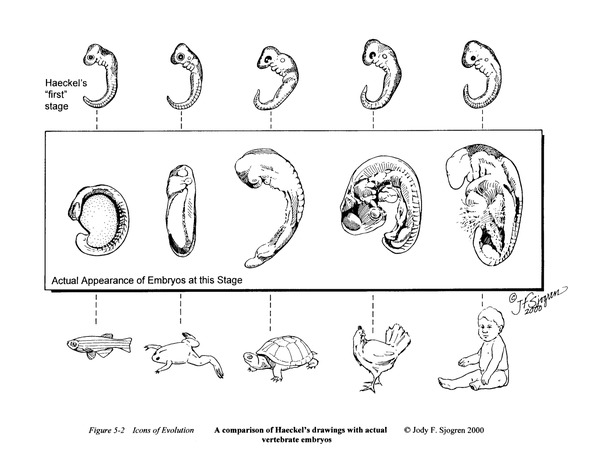
Figure 2: Haeckel’s Embryos
The next icon is Archaeopteryx (Fig. 3). Archaeopteryx was once thought to be a missing link between reptiles and birds. It had teeth and feathers.

Figure 3. Archaeopteryx fossil held by the North Carolina Museum of Natural Sciences, Raleigh, North Carolina. Photo by Dan Reynolds
One problem for it being a forerunner of modern birds is that bird fossils purported to be older than Archaeopteryx have been found. Nevertheless, as recently as 2014, a biology textbook called Archaeopteryx the first bird, and another text published in 2013 refers to Archaeopteryx as “a missing link” between reptiles and birds. Zombie science lives on.
The next icon discussed was industrial melanism of peppered moths. Two species of peppered moths, one light colored, the other dark, lived in England prior to the industrial revolution. The moths lived in trees. Once the industrial revolution was underway, the trunks of the trees were darkened by air pollution. The dark moth variety increased in number relative to the white in the polluted forests. The story goes that the darker moths were better camouflaged by the darker trunks and were less likely to be eaten by predator birds. In the less polluted forests, the lighter colored moths dominated. Pictures of the moths on the tree trunks were taken and published in textbooks. Unfortunately, the pictures were staged with dead moths pinned to the trunks. It is now known that the moths live in the upper canopy and rarely fly during the day. More recent experiments had equivocal results. At best, this would have been a good example of natural selection, the general operation of which is not disputed, but would not have explained the origin of the moths or the color variations. Nevertheless, peppered moths are still held up as “proof of evolution.”
The next icon is Darwin’s Finches (Fig. 4). Observations had shown that finches of the Galapagos Islands with larger beaks were favored in times of drought.
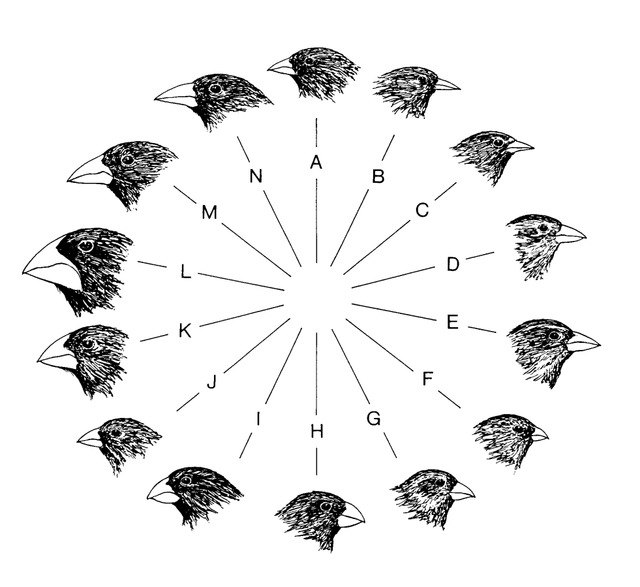
Figure 4. Darwin’s Finches
These birds were better equipped to crack open the thick walled seeds formed during droughts. However, when better weather returned, the finches with smaller beaks reemerged. No new species formed, just reversible changes in overall population composition. Some reports say the number of species has decreased due to hybridization. The various finches on the islands do interbreed, as genetic studies have shown. So there does not appear to be any new species forming at present. As in the case of peppered moths, Darwin’s finches are at best an example of natural selection. What has been observed has nothing to do with macroevolution.
The next icon is Four-winged Fruit Flies (Fig. 5). An artificial mutant fruit fly had four wings. Researchers had selected the mutations. The original balancers behinds the wings on normal flies had been replaced with wings. But the new wings had no muscles and so actually hindered flight and mating. The mutant flies did not survive long in the wild. Discussions of the mutant in recent texts leave out the fact that the flies are crippled. This is not an example of evolution in action.
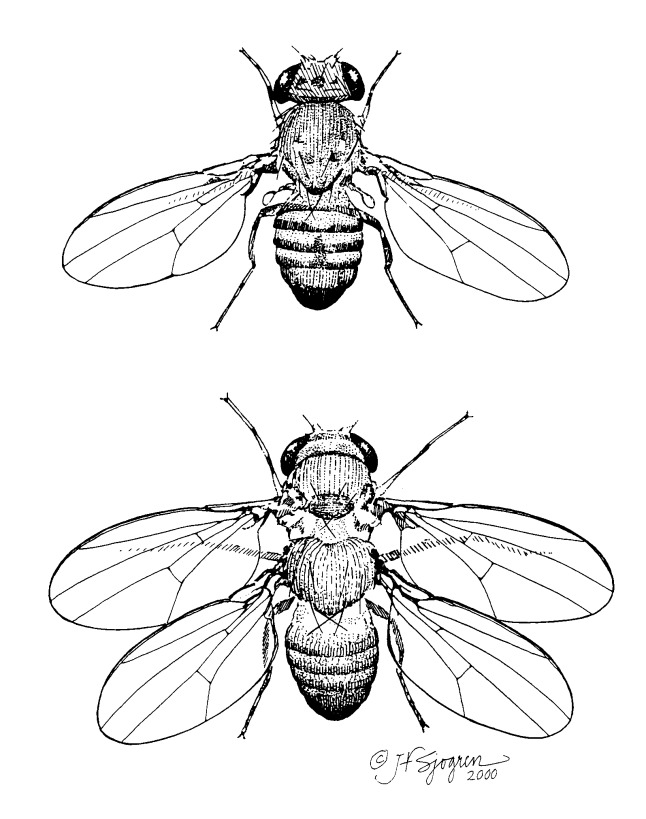
Figure 5. Four-winged Fruit Flies
The last old icon that will be mentioned is the alleged Ape-to-Human evolutionary story (Fig. 6). This is a story of several alleged missing links that have turned out to be either fully human or fully ape. For example, the fossil Ida turned out to be related to lemurs, not humans. Other fossils such as Lucy, Ardi, Sediba, Naledi, and others have only confused the situation. 5
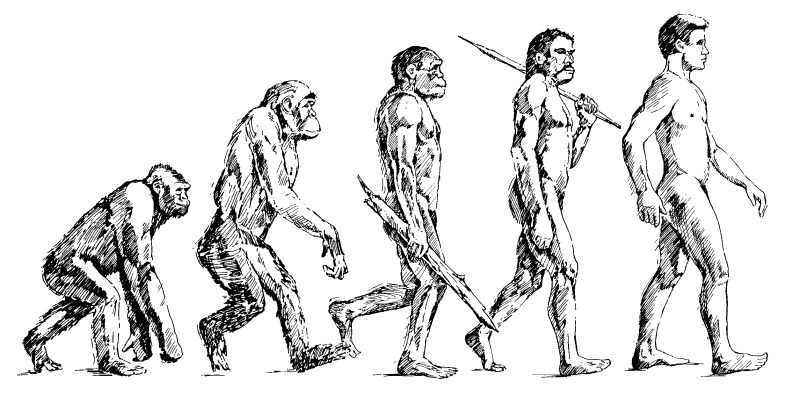
Figure 6. Alleged Evolution of Humans from Apes
Some genetic studies have shown that human and chimp coding DNA are 93.6% similar, not the 99% widely claimed. There are many other problems. 6
Chapter 4: The Secret of Life
Chapter 4: The Secret of Life This is the first chapter where Wells introduces a new icon: The Central Dogma. The Central Dogma of biology is: DNA makes RNA makes proteins makes us. We know that portions of DNA are transcribed into RNA and that the RNA transcripts are translated into proteins in the ribosomes. The dogma assumes that DNA contains the complete blueprint for an organism. Research in the last few decades has unearthed several heritable features of organisms that are not determined by their DNA sequences. Neo-Darwinism assumes that random mutations in DNA acted upon by natural selection is the mechanism of macroevolution. But clearly, if there are heritable characteristics of organisms that are not determined by DNA, then macroevolution (assuming it occurs at all) must depend upon additional factors.
It is well known that different organisms have similar developmental genes. But if different animals have similar developmental genes, why do they develop so differently? Developmental Genetic Regulatory Networks (DGRNs) determine when and where genes are transcribed. Hence animals have similar “toolkit” genes but different “genetic switches.”
In recent years there has been much discussion of epigenetics: factors, other than the base sequences of DNA, that control heritable characteristics. For example, there are heritable characteristics of chromosome structure that don’t depend on the underlying DNA sequence. Epigenetic factors determine when and where genes are transcribed and how cells become committed to specific functions during development. DNA bases can be marked with methyl and acetyl groups signaling when a gene is to be transcribed or ignored.
We now know that a single gene can produce even thousands of proteins. The RNA transcripts of one DNA sequence in fruit flies generates over 18,000 proteins! Hence the original RNA transcript can be spliced/edited in numerous ways before the final messenger RNA (mRNA; produced by transcription of DNA and possibly subsequent RNA editing) transcript is sent to the ribosome. The RNA sequence space available through alternative splicing is facilitated by a molecular machine called the spliceosome. Humans cells are known to use RNA editing via the spliceosome.
Messenger RNA has a “zip code” that determines where it will be delivered in the cell. mRNAs are delivered to the right address along microtubules (“roads”) using molecular “trucks.” The destination address is designated by proteins in the cell membrane. The pattern of address locations in the cell membranes is inherited from ancestor membranes, not from DNA. Hence, the genome does not contain all the information needed to specify an organism.
Organisms don’t start with DNA but with a fertilized egg.
DNA does not explain the three-dimensional development of an embryo. This explains why organisms with similar developmental genes are so different. A higher level of organization is implied. All levels of organization are required to specify an organism. Wells says the various levels of organization that specify an organism are irreducibly complex.
The DGRNs don’t define a cell. The genome/protein/lipid network does. We now know that DNA does not contain the complete blueprint for an organism. Despite the discoveries of epigenetics, the Central Dogma is still taught.
Part 2 of this article will be published in the June 2019 TASC newsletter.
- 1 Wells J (2000) Icons of Evolution: Science or Myth?, Regnery, Washington, DC, 338
- 2Wells J (2017) Zombie Science: More Icons of Evolution, Discovery Institute Press, Seattle, WA, 237
- 3Nested hierarchies are the arrangements of objects based on similarities of properties or features.
- 4Reynolds DW (2017 Jan) Can nature perform the same miracle multiple times? Problems with convergent evolution. TASC Newsletter, https://tasc-creationscience.org/article/can-nature-perform-same-miracle-multipletimes-problems-convergent-evolution Accessed 2019 Apr 15.
- 5See Rupe C, Sanford J (2017) Contested Bones (FMS Publishing) for an in depth discussion of the alleged fossil and genetic evidence for human evolution from an ape ancestor.
- 6Reynolds DW (2015 Apr) On the origin of humans. TASC Newsletter, https://tasc-creationscience.org/article/origin-humans Accessed 2019 Apr 15.